| ||||

 page d'accueil
page d'accueil |  |  | | | ||||||||||||
|
ACTUSCIENCES
| ||||||||||||||||
La réplication du virus du genre influenza |
L’infection commence par l’attachement de l’hémagglutinine à différentes glycoprotéines à la surface de la membrane de la cellule hôte (schéma clé-serrure voir virus sida). Ceci déclenche la fusion des membranes virale et cellulaire par un processus proche de la phagocytose. L’ARN du virus est alors retranscrit (par l’intermédiaire d’une enzyme ARN polymérase virale) en « portion » d’ARN messagers de polarité positive qui serve à la synthèse des protéines virales. La réplication du virus se fait dans le noyau de la cellule. A partir des 8 brins d’ARN constituant le génome du virus, sont synthétisés 10 ARN messagers qui seront traduits dans le cytoplasme en 7 protéines structurales et 3 protéines non structurales. Parallèlement à cela, des protéines virales vont permettre la synthèse de brins d’ARN de polarité négative qui ne seront pas traduits en protéines et qui seront incorporés dans les particules virales filles.
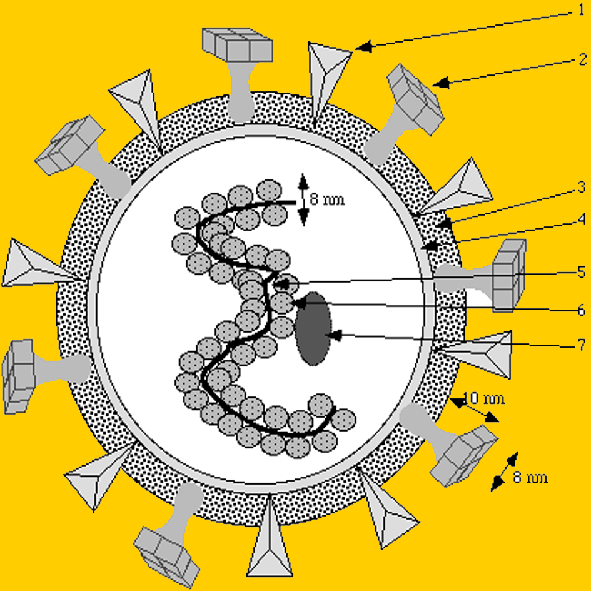
Schéma général du virus influenza : 1. hémagglutinine 2. neuramidase 3. enveloppe 4. couche lipidique 5. RNA segmenté en huit fragments 6. Protéines de la nucléocapside 7. protéine M
Une copie de chacun des 8 fragments d’ARN doit être incorporée dans les virions fils pour qu’ils soient infectieux. On ne sait pas très bien comment le jeu complet de ces segments sont sélectionnés. En considérant une sélection au hasard, la proportion de virus contenant les 8 segments ne serait que de 10%. On considère, généralement, que les virus influenza présentent une grande flexibilité structurale leur permettant d’incorporer plus de 8 segments d’ARN. Certains virus peuvent en contenir jusqu’à 15. Ceci permet d’augmenter la probabilité d’incorporer au moins un jeu complet des 8 segments d’ARN.
Une autre protéine, la protéine M (protéine de matrice), est située en position interne de la membrane virale. Cette protéine sert de support à la construction de la capside du virus qui va progressivement se former et finir par bourgeonner à la surface de la cellule hôte pour donner une nouvelle particule virale libre. Les deux principales protéines membranaires du virus sont très bien connues. L’hémagglutinine possède une extrémité hydrophobe, fixée à l’enveloppe, et une extrémité hydrophile dirigée vers l’extérieur (voir figure). Certaines portions de cette protéine sont conservées entre les différentes générations de virus. Par contre certaines parties sont très variables permettant de distinguer les différents sérotypes viraux. Il en est de même pour la neuraminidase. Elle permet la libération du virus par le détachement des récepteurs de l’hémagglutinine à la surface cellulaire. Lors des différentes épidémies, les souches virales qui sont isolées se caractérisent par les propriétés sérologiques des ces deux protéines. Les variations peuvent être de deux nature :
| |||||||||||||||

 |